Neurophysiology:
Hodgkin Huxley like models
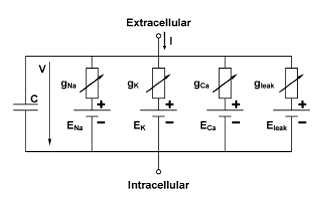
Hodgkin-Huxley like (HH) models simulate neurons and their spiking behaviour close to the biological processes. In the original work of [HODGKIN and HUXLEY 1952] , the spike formation of an action potential is explained by the interaction of the Na and the K ion channel, modelled by different time and voltage dependent conductances, and the capacity of the neurons outer membrane, modelled by a capacitor. This view of a neuron as an electrical circuit is the principle of all HH-models. Newer models however include a variety of different ionic conductances often related to specific ion channels. The basic mathematical description consists of ordinary differential equations of the membranes potential (V) and the activation (m) and inactivation (h) variables of the different ionic conductances.
Hodkin-Huxley models have been used widely to analyse neuronal function in single neurons, neuronal networks, and closed loop conditions, as for instance in the baroflex control of the blood pressure via the Nucleus Tractus Solitarius.
1. HODGKIN, A. L. & HUXLEY, A. F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J Physiol, 1952, 117, 500-544
2. Huguenard, J. R. & McCormick, D. A. Simulation of the currents involved in rhythmic oscillations in thalamic relay neurons. J Neurophysiol, 1992, 68, 1373-1383
3. Rogers, R. F.; Rybak, I. A. & Schwaber, J. S. Computational modeling of the baroreflex arc: nucleus tractus solitarius. Brain Res Bull, 2000, 51, 139-150
4. Rybak, I. A.; Paton, J. F. & Schwaber, J. S. Modeling neural mechanisms for genesis of respiratory rhythm and pattern. I. Models of respiratory neurons. J Neurophysiol, 1997, 77, 1994-2006
5. Schwaber, J. S.; Graves, E. B. & Paton, J. F. Computational modeling of neuronal dynamics for systems analysis: application to neurons of the cardiorespiratory NTS in the rat. Brain Res, 1993, 604, 126-141
6. Vadigepalli, R.; Doyle, F. J. & Schwaber, J. S. Analysis and neuronal modeling of the nonlinear characteristics of a local cardiac reflex in the rat. Neural Comput, 2001, 13, 2239-2271
Signal transduction:
Angiotensin II receptor pathway
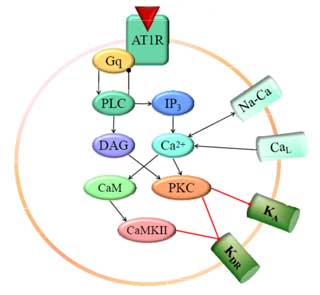
The octapeptide Angiotensin II (AngII) is a multifunctional hormone and is involved in stimulation of water and sodium uptake, vasopressin secretion, increased blood pressure and modulation of baroreflexes.
The cardiovascular actions of Angiotensin II (AngII) within the brain involve increases of vasopressin release, dampening of baroreflexes and stimulation of sympathetic pathways via Angiotensin Type 1 (AT1) receptor activation in the hypothalamus and brain stem, particularly in the Nucleus Tractus Solitarius (NTS). Most of the effects are mediated by the activation of the Angiotensin Type 1 Receptor (AT1R) in the NTS neurons. The significance of the brain AngII system in blood pressure control and hypertension is further supported by studies with spontaneously hypertensive rats (SHR),which show increased AT1 receptor protein [Sun et al. 2003; Sundaram et al. 1997].
The immediate effects of AngII on neuronal activity, that is the inhibition of the firing rate, involve activation of the AT1 receptor and its intracellular signalling pathway [Sumners et al. 2002, 1996]. This includes activation of G-Protein-a subunit and subsequent Phospholipase-C-b (PLC), which rises the concentrations of the second messengers Ca2+ and DAG, activating the Protein-Kinase-C a (PKC) and the Calcium-Calmodulin-dependent Kinase II (CaMKII). PKC reduces the fast activating (Ka) as well as the delayed rectifier (Kdr) potassium current, and activates (probably through inhibition of a negative influence of G-bg subunit) the total transmembrane calcium current [Pan et al. 2001; Zhu et al. 1997]. Kdr is further reduced by CaMKII [Zhu et al. 1999]. The potassium channel subtypes involved in this inhibition are Kv2.2 for the Kdr and Kv1.4 for the Ka respectively [Pan et al. 2000; Gelband et al. 1999]. Their modulation is most probably the effect of direct phosphorylation through the above mentioned kinases.
Longer term actions of the AT1R pathway involve Janus kinase 2 (JAK2) association in a G-protein independent manner and the subsequent activation of transcription (STAT) molecules, as well as transactivation of tyrosine kinase pathways (e.g. Ras, MAPK), which involves the transcription factors c-Jun, c-Fos and NF-kappaB [Sumners et al. 2002; Richards et al. 1999; Huang et al. 1998; Lu et al. 1998; Marrero et al. 1996].
1. Gelband, C. H.; Warth, J. D.; Mason, H. S.; Zhu, M.; Moore, J. M.; Kenyon, J. L.; Horowitz, B. & Sumners, C. Angiotensin II type 1 receptor-mediated inhibition of K+ channel subunit kv2.2 in brain stem and hypothalamic neurons. Circ Res, 1999, 84, 352-359
2. Huang, X. C.; Deng, T. & Sumners, C. Angiotensin II stimulates activation of Fos-regulating kinase and c-Jun NH2-terminal kinase in neuronal cultures from rat brain. Endocrinology, 1998, 139, 245-251
3. Marrero, M. B.; Paxton, W. G.; Schieffer, B.; Ling, B. N. & Bernstein, K. E. Angiotensin II signalling events mediated by tyrosine phosphorylation. Cell Signal, 1996, 8, 21-26
4. Pan, S.; Sumners, C. & Gelband, C. Kv1.4 underlies angiotensin II-mediated inhibition of neuronal A-type K+ current Biophys J, 2000, 78(1), 450A-450A
5. Pan, S. J.; Zhu, M.; Raizada, M. K.; Sumners, C. & Gelband, C. H. ANG II-mediated inhibition of neuronal delayed rectifier K+ current: role of protein kinase C-alpha. Am J Physiol Cell Physiol, 2001, 281, C17-C23
6. Richards, E. M.; Raizada, M. K.; Gelband, C. H. & Sumners, C. Angiotensin II type 1 receptor-modulated signaling pathways in neurons. Mol Neurobiol, 1999, 19, 25-41
7. Sumners, C.; Fleegal, M. A. & Zhu, M. Angiotensin AT1 receptor signalling pathways in neurons. Clin Exp Pharmacol Physiol, 2002, 29, 483-490
8. Sumners, C.; Zhu, M.; Gelband, C. H. & Posner, P. Angiotensin II type 1 receptor modulation of neuronal K+ and Ca2+ currents: intracellular mechanisms. Am J Physiol, 1996, 271, C154-C163
9. Sun, C.; Du, J.; Sumners, C. & Raizada, M. K. PI3-kinase inhibitors abolish the enhanced chronotropic effects of angiotensin II in spontaneously hypertensive rat brain neurons. J Neurophysiol, 2003, 90, 3155-3160
10. Sundaram, K.; Johnson, S. M. & Felder, R. B. Altered expression of delayed excitation in medial NTS neurons of spontaneously hypertensive rats. Neurosci Lett, 1997, 225, 205-209
11. Zhu, M.; Neubig, R. R.; Wade, S. M.; Posner, P.; Gelband, C. H. & Sumners, C. Modulation of K+ and Ca2+ currents in cultured neurons by an angiotensin II type 1a receptor peptide. Am J Physiol, 1997, 273, C1040-C1048
Systems theory:
Model reduction
First principle models derived from some physical/chemical law (as for instance the law of mass action) become easily unsuitable large, especially in cellular reaction networks. This makes the analysis challenging and the gaining of understanding from complex models may be nearly as difficult as from the original system. A model should therefore, on the one hand, be as simple as possible, but on the other explain the phenomenon under investigation as realistic as possible. The aim of model reduction is to achieve the simplification of the model without loosing the essential parts and interconnections of the system.
Nonlinear dynamics
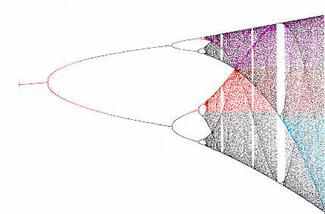
Models arising from a biological background are mostly non-linear in nature. A linear analysis of these models is only locally valid, and in some cases not applicable at all. For a general nonlinear scheme, only very restricted statements can be drawn, which makes the simulation an important part of the systems analysis. Bifurcation analysis for instance tackles the question of the number of steady-states, limit cycles and their stability properties.
However, in order to develop suitable analysis methods, one has often to consider specific classes of models. Reaction kinetic models following the law of mass action is an example for such a model class. Feinberg, Horn and Jackson have been able to obtain results that are globally valid by transferring the non-linear problem to a problem of linear algebra.
Observability and observer design
Often, not all variables of interest can be measured directly. Given a dynamic model of the system, it is of particular interest whether the not measured or not measurable states can be inferred from the measurements.
Especially in cell biological systems, the measurement possibilities a very limited. Often, only a few measured variables at a few time-points are available, which are in addition superposed with noise. This and the complexity and nonlinearity of cell biological systems demands extensive requirements on the observer and the observer design.